
testicoli
I testicoli [ Hodn ] o (più raramente) il o Hode [ hoːdə ] (tramite il medio alto tedesco Hode e il vecchio alto tedesco hodo di Indo - * skeu (t) o "coprire, coprire") testicolo (dal latino testiculus , diminutivo di testis "testimone [di virilità ], testicolo", plurale: testicoli ; greco antico ὄρχις orchis ), tecnicamente anche testicolo e testicolo , è un organo sessuale maschile interno accoppiato di molti animali tissutali che si riproducono sessualmente . Come le ovaie degli individui femminili, appartiene alle gonadi ( gonadi ) e produce i filamenti seminali ( sperma ). Inoltre, nei testicoli si formano gli ormoni sessuali maschili ( androgeni ), in particolare il testosterone . Nei vertebrati, i testicoli si formano embrionale nella cavità addominale, ma nella maggior parte dei mammiferi migrano nello scroto (lo scroto ).

anatomia
Mammiferi
Dimensioni e posizione

Il testicolo umano è grosso modo a forma di prugna , pesa circa 20 grammi ed ha un volume medio di 20-25 ml, la lunghezza media è di 5 cm, lo spessore di circa 3 cm. I testicoli non si sviluppano alla loro piena dimensione fino alla pubertà e raggiungono la loro dimensione massima nella 4a decade di vita. Con l'età, la dimensione del testicolo diminuisce di nuovo. Il volume del testicolo fornisce informazioni sulle condizioni funzionali del testicolo, tra le altre cose. Se il volume del testicolo è inferiore a 8 ml, si può presumere che la produzione di sperma funzioni solo in misura limitata o non funziona affatto. Il testosterone, invece, a volte viene ancora prodotto fino a un volume di 1,5 ml; al di sotto di questo, il testicolo è solitamente disfunzionale.
Nei mammiferi , la forma del testicolo varia da arrotondata a ovoidale. Ci sono chiare differenze di dimensioni, ma non c'è una stretta relazione con il peso corporeo. I testicoli più grandi del mondo animale hanno balene franche australi , con ogni 500 kg costituiscono il 2% del peso corporeo. Roditori , pecore (fino a 300 g ciascuno) e maiali domestici (fino a 750 g ciascuno) hanno testicoli relativamente grandi, mentre i predatori sono relativamente piccoli . Negli animali con un periodo di riproduzione stagionale, anche la dimensione del testicolo è soggetta a fluttuazioni stagionali, i testicoli sono significativamente più grandi nella stagione degli amori rispetto al periodo di dormienza.
Nella maggior parte dei mammiferi, entrambi i testicoli si trovano in individui sessualmente maturi nello scroto ( scroto ) o in borse di pelle simili allo scroto. I testicoli sorgono nella cavità addominale , ma migrano attraverso il canale inguinale nello scroto intorno al momento della nascita, nei roditori non fino alla pubertà . Questo processo è chiamato discesa testicolare ( Descensus testis ). In alcuni mammiferi ( ad esempio criceti , pipistrelli ) c'è una discesa testicolare stagionale e i testicoli sono solo al di fuori della cavità addominale durante la stagione degli amori. All'interno dei mammiferi, invece, esistono alcuni gruppi di animali in cui i testicoli rimangono generalmente nella cavità addominale, i cosiddetti testiconda . I testicoli possono rimanere nella sede del sistema (come negli elefanti ) oppure possono discendere, ma indugiare ancora nella cavità addominale (ad esempio nelle balene , vedi anche tabella).
| Testiconda | ||
| Nessuna discesa del testicolo | Discesa incompleta del testicolo | Discesa testicolare stagionale |
| Monotremi , chrysochloridae , toporagni , Igeltenreks , iraci , elefanti , trichechi , bradipo tridattilo , formichieri | Balene , armadilli | Talpe , punteruoli , oritteropi , toporagni , pipistrelli , alcuni roditori |
Struttura anatomica
La struttura anatomica esterna del testicolo si basa sull'epididimo che confina e si fonde con esso . La sezione del testicolo che punta alla testa dell'epididimo è chiamata estremità della testa ( Extremitas capitata ), la sezione che punta alla coda dell'epididimo è chiamata estremità della coda ( Extremitas caudata ). All'estremità della coda c'è spesso un rudimento senza funzione, a forma di verruca, del cosiddetto dotto di Müller , noto come appendice testicolare ( appendice testis , un'idatide di Morgagni ). Il bordo rivolto verso l'epididimo è il bordo dell'epididimo ( Margo epididymalis ), opposto è il bordo libero ( Margo liber ). Inoltre, si possono distinguere una superficie che punta verso il centro ( Facies medialis ) e una superficie che punta verso l'esterno ( Facies lateralis ).
I testicoli scendono in un rigonfiamento nel peritoneo e nella fascia interna del tronco (qui denominata Fascia spermatica interna ), il processus vaginalis . Il processo della vagina è una delle guaine testicolari all'interno dello scroto. La parte del peritoneo di questa protuberanza è chiamata pelle vaginale ( tunica vaginalis testis ). Riveste l'interno dello scroto (cosiddetto lenzuolo murario, lamina parietalis o periorchium ), quindi si trasforma all'interno come una doppia lamella e ricopre i testicoli come un lenzuolo intestinale ( lamina visceralis o epiorchium ). C'è uno spazio molto stretto tra le due foglie, il cavum vaginale , che fa sì che il testicolo possa muoversi nello scroto. Il punto di connessione tra le due foglie è il testicolo crosus ( mesorchium ), che serve per attaccare il testicolo allo scroto. Il testicolo è anche all'estremità della coda con una breve striscia collegata all'epididimo (banda del testicolo , legamento del testicolo proprio ). Questo continua dalla coda dell'epididimo come legamento della coda dell'epididimo ( legamento caudae epididimidis ) e inoltre attacca i testicoli indirettamente al pavimento dello scroto. Il muscolo di sollevamento dello scroto ( Musculus cremaster ) si attacca anche al processo della vagina, poiché un dispositivo protettivo avvicina i testicoli alla parete addominale quando viene toccato o freddo. Nei roditori e nei mammiferi con discendenza testicolare stagionale, raramente anche nei singoli individui di altri mammiferi, il muscolo può riportare completamente il testicolo nella cavità addominale (" testicolo pendolo ").
Una spessa capsula di tessuto connettivo biancastro , la tunica albuginea, si trova direttamente sotto la copertura del peritoneo del testicolo . Assicura la resistenza meccanica dell'organo e mantiene una certa pressione interna. Da questa capsula i setti tirano verso l'interno e suddividono i testicoli in lobuli testicolari ( lobules testis ). Il testicolo dell'uomo ha circa 350 lobuli testicolari. I setti formano anche un corpo di tessuto connettivo, il mediastino testicolo , che nell'anatomia umana è anche chiamato corpus highmori .
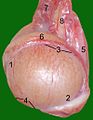
Testicoli, epididimo e funicolo spermatico di un gatto maschio :
1 estremità della testa
2 estremità della coda
3
bordo dell'epididimo 4 bordo libero
5 mesentere testicolare
6 epididimo
7 plesso dell'arteria e della vena testicolare
8 dotto spermatico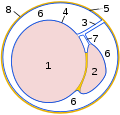
Sezione trasversale del processo vaginale:
1 testicolo
2 epididimo
3
mesentere testicolare 4 foglio d'organo della membrana vaginale
( epiorchio )
5 foglio di parete della pelle vaginale
( periorchio )
6 cavo vaginale
7 mesentere epididimale
8 fascia spermatica interna
Sezione attraverso un testicolo di toro:
(vasi sanguigni iniettati con gelatina rossa )
1 parenchima testicolare
2 mediastino testicolo
3 tunica albuginea
4 coda
dell'epididimo 5 testa dell'epididimo
6 funicolo spermatico con
convoluzione del viticcio dell'arteria testicolare



Vasi e nervi
.jpg)
Il testicolo viene rifornito di sangue attraverso l'arteria testicolare ( arteria testicolare ). Nasce, corrispondente alla posizione dell'attacco embrionale del testicolo (vedi sotto), direttamente dietro l' arteria renale, direttamente dall'aorta addominale nella regione lombare. Negli animali con discesa testicolare, l'arteria testicolare deve allungarsi di conseguenza e corre lungo la parte posteriore della parete addominale in un breve mesentere ( mesorchio prossimale ) fino al canale inguinale . Fuori dalla cavità addominale, entra nel cordone spermatico . Qui giace in stretti avvolgimenti a spirale, la cosiddetta convoluzione a viticcio . Nei tori , ad esempio, un tratto di arteria lunga due metri è alloggiato su un cordone spermatico lungo 13 cm. Il fascio di viticci è avvolto nel plesso viticcio ( Plexus pampiniformis ) della vena testicolare ( Vena testicularis ). Questo crea un'ampia area di contatto tra il sangue in entrata e in uscita, che funge da scambiatore di calore . La temperatura nello scroto è di qualche grado inferiore alla temperatura corporea interna, essenziale per la formazione di spermatozoi fertili nei mammiferi con discendenza testicolare. Il sangue in entrata nell'arteria viene raffreddato da questa disposizione dal sangue in uscita dalla vena.
L'arteria testicolare corre lungo il bordo dell'epididimo fino alla fine della coda del testicolo. Da lì si sposta all'interno della capsula testicolare all'estremità libera nella maggior parte dei mammiferi (i ruminanti , ad esempio, fanno un'eccezione ) fino all'estremità della testa. I suoi rami si snodano nella tunica albuginea sulle superfici laterali e passano sui setti testicolari interni al mediastino testicolo e da lì centrifugamente tornano ai tubuli seminiferi, attorno ai quali formano una rete capillare .
L' innervazione del testicolo è mediata dal simpatico , parte del sistema nervoso autonomo . Le fibre nervose provengono dal tronco della regione lombare e, avvolgendosi come una treccia intorno all'arteria testicolare ( plesso testicolare , sinonimo: nervo spermatico superiore ), si estendono ai testicoli. Un secondo gruppo di fibre nervose va dai gangli trasversali del tronco con il dotto spermatico ( dotto deferente ) ai testicoli ( plesso deferenziale , sin. nervo spermatico inferiore ). Le fibre nervose efferenti raggiungono principalmente i vasi sanguigni e regolano così il flusso sanguigno e la temperatura del testicolo. Attualmente è in discussione la partecipazione al controllo fine della formazione degli spermatozoi , del trasporto degli spermatozoi e della produzione di ormoni nei testicoli di alcuni mammiferi, ma questo controllo viene effettuato principalmente tramite gli ormoni . I corpi cellulari di quelle fibre nervose che portano informazioni al sistema nervoso centrale ( visceroaffezioni ) si trovano nei gangli delle radici dorsali della regione lombare. Trasmettono sensazioni di dolore ( dolore intestinale), ma gran parte dell'elevata sensibilità del testicolo al dolore viene veicolata attraverso le fibre nervose sensibili delle guaine testicolari (rami del nervo genitofemorale ). L'elevata sensibilità agli stimoli tattili rende i testicoli e lo scroto una zona erogena . D'altra parte, la grande sensibilità al dolore viene sfruttata anche nel BDSM ( tortura del cazzo e della palla ) e nella tortura ; La compressione dei testicoli può provocare uno shock . Una nevralgia del nervo genitofemorale, che può verificarsi, ad esempio, dopo la correzione chirurgica di un'ernia inguinale , si manifesta nel dolore ai testicoli.
I vasi linfatici del testicolo corrono insieme alle vene testicolari ai linfonodi lombari ( Lymphonodi lumbales ) posteriormente all'aorta, nei mammiferi domestici anche ai linfonodi iliaci ( Lymphonodi iliaci mediales ) al ramo aortico. Nel cancro ai testicoli, possono verificarsi metastasi in questi linfonodi , che si trovano nello spazio retroperitoneale dell'addome .
Cordati rimanenti
In tutti gli altri cordati , i testicoli si trovano nella cavità del corpo e sotto i reni.
Nel skullless non c'è testicolo compatto, le gonadi sono ancora segmentati, in Asymmetron solo testicolo destro è presente. Nella missina , anche il testicolo allungato è formato solo da un lato a forma di banda lobata. Le bocche rotonde hanno testicoli accoppiati che si estendono per l'intera lunghezza della cavità corporea.
All'interno del pesce cartilagineo , solo nello squalo collare , lo squalo anatomicamente e morfologicamente più primitivo, l'intero sistema testicolare si sviluppa in una gonade allungata, negli altri rappresentanti, come negli altri vertebrati , solo la sezione mediana (vedi sotto). I testicoli i veri squali sono allungate, e quelli dei raggi sono corto e piatto. Nel palombo e nei raggi elettrici , i testicoli sono molto avanti, vicino al pericardio , altrimenti nella sezione centrale o posteriore del tronco. Nella maggior parte dei pesci ossei , i testicoli sono allungati. Nel vero pesce osseo , si trovano sotto i reni e la vescica natatoria e sono attaccati tramite un mesorchio. In alcune specie di pesce persico , entrambi i testicoli sono fusi insieme all'estremità posteriore. La sottile tunica albuginea è occasionalmente pigmentata nei pesci ossei . Allo stesso tempo, il tessuto ovarico può apparire nei testicoli dei pesci (ermafrodita, ovotestis ). Il branzino e l' orata sono ermafroditi , quindi hanno sia i testicoli che le ovaie e possono cambiare sesso nel corso della loro vita.
Gli anfibi hanno testicoli allungati ( anfibi coda , anfibi striscianti ) o arrotondati ( anfibi rana ). Sono attaccati alla parete del tronco o al rene dell'urna tramite un mesorchio . Un corpo grasso distinto è formato medialmente (nella direzione del piano mediano ) del testicolo. Nelle salamandre , diversi reparti testicolari sono fusi in un lobo, il numero di reparti aumenta con l'età. Negli anfibi, le vie urinarie e seminali iniziano a separarsi. I dotti epididimali ( ductuli epididymidis ) si aprono nel dotto di Wolff solo poco prima della cloaca . I rospi maschi sono ermafroditi. Davanti ai testicoli si trova l' organo dell'offerente , un ovaio primitivo, che si sviluppa dall'anlage gonadico anteriore .
Nei rettili , i testicoli si trovano davanti e sotto i reni in prossimità delle ghiandole surrenali . I testicoli sono ovali, più arrotondati nelle tartarughe e allungati nei serpenti . Gli epididimi sono mediali (verso il centro del corpo) del rispettivo testicolo. La struttura del tessuto connettivo del testicolo è poco sviluppata.
Anche negli uccelli i testicoli si trovano davanti ai reni sulle ghiandole surrenali. A differenza dell'ovaio, che negli uccelli è solo unilaterale, i testicoli sono sempre accoppiati. La dimensione del testicolo mostra le maggiori fluttuazioni stagionali all'interno dei vertebrati; aumenta, ad esempio, nei passeriformi fino a 800 volte durante la stagione degli amori e diminuisce nuovamente durante la dormienza dell'accoppiamento. In un maschio di casa i testicoli sono nella stagione degli amori lunghi circa 8 cm e larghi 4,5 cm. La struttura del tessuto connettivo del testicolo è solo leggermente sviluppata negli uccelli, la tunica albuginea è sottile e non c'è mediastino testicolo . L' esame endoscopico del testicolo giocava un ruolo importante nella determinazione del sesso delle specie che non mostrano dimorfismo sessuale , ma oggi è stato ampiamente sostituito da metodi di biologia molecolare .
invertebrati
La riproduzione asessuale e sessuale avviene all'interno degli cnidari . Durante la riproduzione sessuale, le cellule germinali vengono rilasciate nell'acqua circostante o nello spazio gastrico facendo esplodere l' epitelio . Le meduse costali sono generalmente ermafroditi e hanno testicoli e ovaie incorporati nella mesogloea sotto le piastrine a pettine ("costole") .
Bocche primordiali (protostomia)
La maggior parte dei platelminti (piastrine) sono ermafroditi. Non hanno una cavità corporea ( celoma ), i testicoli giacciono adiacenti alle ovaie in uno spazio di tessuto connettivo ( interstizio ) all'interno dell'animale, nel caso di tenie all'interno di ciascun arto di tenia ( proglottide ). Alcuni platelminti ( Catenulida e Acoelomorpha ) non hanno gonadi. Anche a pelo ventre (Gastrotricha) non hanno celoma, le specie d'acqua dolce si riproducono asessualmente per partenogenesi . I rappresentanti dell'acqua salata sono ermafroditi, ma solo le gonadi di un sesso sono funzionalmente attive. Le cellule germinali vengono rilasciate tramite pile di sperma . I rotiferi (Rotatoria) hanno uno pseudoceloma con testicoli appaiati o non appaiati, in alcune specie sono presenti i cosiddetti maschi nani , in altre no maschi, la riproduzione avviene poi tramite generazione vergine. I ciclofori si riproducono asessualmente nella cosiddetta fase di alimentazione. Nella riproduzione sessuale si verificano maschi nani. Hanno due "testicoli" esterni e un organo copulatore vicino al disco adesivo. Nel caso dei vermi graffianti (Acanthocephala), un cosiddetto legamento genitale attraversa il corpo, nel terzo inferiore del quale si trovano i due testicoli. Un condotto spermatico va da ciascun testicolo al pene.
Priapulida e animali da corsetto (Loricifera) hanno testicoli appaiati che si uniscono ai protonefridi per formare un apparato urinario e sessuale . Nei nematodi (Nematoda) ci sono sia specie di sesso separato che ermafroditi. Il testicolo allungato si trova sotto l'intestino e si apre anche in esso (vedi fig.). I vermi filiformi (Nematomorpha) hanno testicoli appaiati nello pseudoceloma. I tardigradi (Tardigrada) sono sessi separati. Mentre la cavità corporea rappresenta uno pseudoceloma, c'è un vero e proprio celoma attorno ai testicoli spaiati. Colibus (Onychophora) hanno testicoli accoppiati che si aprono in un comune dotto deferente attraverso i dotti. Negli artropodi (Arthropoda: millepiedi , insetti , crostacei e chelicerati ) i maschi hanno testicoli appaiati nello pseudocele ( emocele ) dell'addome o nei segmenti posteriori del tronco. Ci sono anche ermafroditi in alcuni artropodi.
Gli anellidi hanno testicoli appaiati. I vermi della cintura sono per lo più ermafroditi, ma si fertilizzano a vicenda. I testicoli si trovano nel celoma , nei lombrichi nel 10° e nell'11° segmento del corpo e i dotti deferenti si aprono verso l'esterno nel 15° segmento. Le multi-setole , d'altra parte, sono per lo più unisessuali e i rappresentanti maschili hanno testicoli in ogni segmento del corpo. Nei molluschi (Mollusca) sono presenti ermafroditi, gonadi ermafrodite ( Ovotestis ) e forme sessuali separate. Il celoma è ridotto a due cavità attorno alle gonadi ( gonadocele ) e attorno al cuore ( pericardio ) dietro di esso . Negli armpods (Brachiopoda), i testicoli giacciono nel celoma (più precisamente nel metacoel ), i gameti derivano tramite i metanephridia .
Neumünder (Deuterostomia)
Negli echinodermi (Echinodermata), le balene di mare hanno un solo testicolo, nei ricci di mare e nelle balene di mare i testicoli riempiono quasi l'intero metacele, nei gigli di mare (da tre a cinque gonadi) e nelle stelle marine (due per braccio) sdraiarsi tra le braccia e aprire tra di loro Braccia con un'apertura genitale ciascuna.

Organi interni di un pesce osseo:
7 testicoli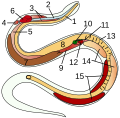
Organi interni di un serpente:
14 testicoli
Sito di un gallo domestico:
(esclusi gli organi digestivi)
3 testicoli
Ascaridi:
5 testicoli
Progetto di un baccello con gonadi nel metacele




Struttura fine e funzione nei vertebrati
I lobuli testicolari contengono ciascuno da due a quattro tubuli seminiferi arrotolati ( tubuli seminiferi contorti s. Convoluti ), che rappresentano il parenchima testicolare . Sono lunghi da 50 a 80 cm e hanno un diametro da 150 a 300 µm. La sua parete è costituita da un guscio di tessuto connettivo con miofibroblasti contrattili , una membrana basale e l'epitelio germinale ( epithelium spermatogenicum ).
Questo epitelio è costituito da spermatozoi o cellule germinali ( Cellulae spermatogenicae ) e cellule del Sertoli . Gli spermatozoi sono formati dalle cellule germinali ( spermatogenesi ). Poiché la spermatogenesi è il compito più importante del testicolo, le cellule germinali sono anche più frequentemente presenti nei testicoli in termini di quantità. Durante la formazione degli spermatozoi, i successivi stadi di sviluppo delle cellule germinali ( spermatogoni → spermatociti → spermatidi → spermatozoi) vengono trasportati gradualmente verso il lume . La formazione dello sperma richiede tra 35 (topo, maiale) e 64 giorni (umano), ma è necessaria un'ulteriore maturazione nell'epididimo affinché lo sperma possa essere fecondato. Questo dura una settimana nella maggior parte dei mammiferi e da 8 a 17 giorni nell'uomo. Negli uomini, circa 200-300 milioni di spermatozoi vengono rilasciati dall'epididimo per eiaculazione . Con l'eiaculazione più frequente, la quantità di sperma diminuisce perché la capacità di produzione giornaliera di sperma è limitata. Dipende dalla massa del testicolo e dal numero di cellule del Sertoli e negli uomini è compreso tra 45 e 200 milioni di spermatozoi al giorno.
Il secondo importante componente dei tubuli seminiferi sono le cellule del Sertoli ( Epitheliocyti sustentantes ). Sono lunghi circa 70-80 µm e corrono radialmente attraverso l'intero epitelio germinativo fino al lume. Le cellule del Sertoli hanno una funzione di sostegno e nutrimento per gli spermatozoi, nutrono gli spermatozoi, assicurano il loro corretto ambiente ormonale e utilizzano i movimenti del plasma per trasportarli al lume. Inoltre, le cellule del Sertoli fagocitano spermatozoi degenerati e detriti cellulari che si formano durante lo sviluppo degli spermatozoi. Le cellule del Sertoli sono attivate dall'ormone follicolo-stimolante dell'ipofisi (FSH) controllato, la sua distribuzione sulla formazione dell'ormone inibina B influenza. Inoltre, secernono la proteina legante gli androgeni , l' ormone anti- Müller e un liquido seminale ricco di potassio .
Le cellule del Sertoli hanno numerosi processi che circondano le cellule germinali. Questi processi cellulari si collegano alla base dei tubuli seminiferi tramite giunzioni strette con quelli delle cellule del Sertoli limitrofe e formano così la cosiddetta barriera emato-testicolare . Questo termine è in realtà fuorviante, perché questa barriera non si trova tra il sangue e il tessuto testicolare, ma corre tra gli spermatogoni e gli spermatociti, dividendo così i tubuli testicolari in modo circolare in un compartimento basale e uno rivolto verso il lume (adluminale) . La barriera sangue-testicoli è impermeabile alla maggior parte delle proteine e protegge lo sperma dai mutageni e dalle difese dell'organismo . Quest'ultimo è necessario perché i primi spermatozoi emergono solo dopo che i linfociti sono stati impressi (vedi autotolleranza ), quindi il sistema immunitario li considererebbe cellule estranee. Tuttavia, anche le citochine antinfiammatorie , il cui rilascio è presumibilmente androgeno- dipendente, e le cellule del sistema immunitario nei testicoli ( cellule dendritiche , macrofagi ) svolgono un ruolo protettivo contro le reazioni autoimmuni .
In molti vertebrati, i tubuli seminiferi contorti si fondono in un breve tubulo seminifero rettilineo ( tubulus seminifer rectus ) ad entrambe le estremità . I tubuli diritti sono rivestiti da un epitelio monostrato e si aprono in un sistema di canali nel mediastino, la rete testicolare ( rete testis ). Anche il sistema canalico della rete testicolare è per lo più rivestito da un epitelio a strato singolo (a due strati nei bovini). Negli uomini, stalloni e roditori, invece , la rete testicolare è prevalentemente alla periferia dei testicoli ("rete extratesticolare"). Diversi meandri ductuli efferentes testis corrono dalla rete testicolare nella testa dell'epididimo e si uniscono lì per formare il dotto epididimale. Nei mammiferi sono presenti circa 15 duttuli efferenti, il numero varia all'interno dei vertebrati tra uno (es. raggi ) e 32 (es. axolotl ).
Il tessuto tra i tubuli seminiferi contorti è chiamato interstizio . Nella maggior parte dei vertebrati, costituisce circa il 10-20% del tessuto testicolare, in casi estremi come la marmotta quasi il 70%. Oltre al tessuto connettivo, ai vasi sanguigni e alle fibre nervose , l' interstizio contiene anche cellule di Leydig ( Endocrinocyti interstitiales ). Tramite speciali contatti cellulari ( giunti gap ), formano associazioni cellulari interconnesse, i cosiddetti sincizi funzionali . A seconda dell'ormone luteinizzante (LH), le cellule di Leydig producono ormoni sessuali maschili (androgeni come testosterone e androstanolone ) e ossitocina , che favorisce la motilità dei tubuli seminiferi. Il testicolo è quindi anche un organo endocrino . Il testosterone fa maturare gli spermatidi nei tubuli seminiferi. Per passare attraverso la barriera emato-testicolare al suo luogo d'azione, ha bisogno della proteina legante gli androgeni delle cellule del Sertoli. Gli androgeni hanno anche una varietà di effetti nel corpo, tra cui promuovere lo sviluppo di caratteristiche sessuali secondarie , avere un effetto anabolico e controllare il comportamento sessuale . Inoltre, le cellule di Leydig producono numerosi altri peptidi ormonalmente attivi che agiscono sulle cellule vicine ( paracrine ) o sulla stessa cellula di Leydig ( autocrine ) che le compongono .

Quadro istologico del parenchima testicolare di un verro:
1 lume di un tubulo seminifero contorto
2 spermatidi
3 spermatociti
4 spermatogoni
5 cellule di Sertoli
6 miofibroblasti
7 cellule di Leydig
8 capillari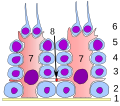
Epitelio germinale del tubulo seminifero :
1 membrana basale
2 spermatogoni
3 spermatociti 1° ordine
4 spermatociti 2° ordine
5 spermatidi
6 spermatidi maturi
7 cellula del Sertoli
8 giunzione stretta
(barriera sangue-testicolo)
Schema della struttura interna del testicolo e dell'epididimo :
1 tunica albuginea
2 setti
3 lobi testicolari
4 mediastino testicolo
5 tubuli seminiferi contorti
6 tubuli seminiferi diritti
7 rete testis
8 ductuli efferentes testis
9 epididimo
10 inizio del dotto deferente



Controllo ormonale

Il controllo ormonale del testicolo avviene a scatti, da cellule nervose nel eminenza mediana nella ipotalamo formata gonadoliberin (GnRH). Il GnRH non agisce direttamente sui testicoli, ma stimola la formazione degli ormoni LH e FSH nella ghiandola pituitaria anteriore . Il rilascio di questi ormoni è controllato anche dai testicoli stessi tramite un meccanismo di feedback negativo: la secrezione di FSH è inibita dall'inibina B prodotta dalle cellule del Sertoli, mentre la secrezione di GnRH è inibita dal testosterone prodotto dalle cellule di Leydig . Le fluttuazioni stagionali delle dimensioni e dell'attività dei testicoli in molti animali sono mediate dalla soppressione della secrezione di GnRH durante il riposo sotto l'influenza della lunghezza della luce del giorno. L'esatto meccanismo non è ancora noto nei dettagli: nei mammiferi, gli oppioidi , i neuroni dopaminergici e la melatonina sono presumibilmente coinvolti in questo ciclo di controllo, negli uccelli anche gli ormoni tiroidei .
LH si lega a un recettore di membrana delle cellule di Leydig e quindi induce la sintesi degli androgeni . In questo caso sono possibili gradualmente colesterolo , tra l'altro, pregnenolone e progesterone , convertiti in testosterone, utilizzando due diverse vie sintetiche (Δ4 e Δ5). L'effetto LH sulle cellule di Leydig è potenziato dalla prolattina , ma con una sovrapproduzione di prolattina si verifica una down-regolazione dei recettori LH per inibire la sintesi del testosterone. LH può anche indurre la formazione di androgeni nella corteccia surrenale ; il deidroepiandrosterone ivi formato raggiunge i testicoli attraverso il sangue e può essere utilizzato come precursore del testosterone. Circa il 97% degli androgeni si forma nei testicoli (circa 7 mg/die negli uomini), la parte restante nelle ghiandole surrenali . Gli androgeni agiscono sull'epitelio germinale e, legati a una proteina, vengono trasportati anche agli altri organi bersaglio attraverso il sangue.
L'FSH si lega ai corrispondenti recettori sulle cellule del Sertoli. Sia l'FSH che il testosterone controllano la spermogenesi. L'FSH avvia la spermiogenesi, il testosterone promuove la divisione cellulare mitotica e meiotica e quindi la formazione di spermatociti dagli spermatogoni, mentre l'FSH a sua volta provoca la maturazione finale degli spermatidi in sperma.
La soppressione della stimolazione ormonale della funzione testicolare è attualmente oggetto di intensa ricerca nello sviluppo di contraccettivi per gli uomini . Il testosterone o la sua combinazione con antagonisti del GnRH o gestageni come il progestinico sono attualmente considerati i candidati più promettenti. Conducono a concentrazioni di testosterone notevolmente ridotte all'interno del testicolo e quindi a una riduzione forte o completa della formazione degli spermatozoi. In medicina veterinaria, una preparazione sulla base del GnRH analogica Deslorelin ( Suprelorin ® ) è stato approvato dal 2008 , che sopprime la fertilità nei maschi per diversi mesi. Inoltre, è approvato un vaccino per suini ( Improvac ® ) che porta alla formazione di anticorpi contro il GnRH e quindi sopprime la funzione testicolare.
Storia dello sviluppo

Il presupposto per la riproduzione sessuale è la separazione delle cellule specializzate per la riproduzione (cellule germinali) dalle cellule normali del corpo (cellule somatiche). Questa separazione è già effettuata nei ciliati sotto forma di micronucleo , più chiaramente che nelle alghe sferiche , dove il gruppo cellulare principale delle cellule del corpo si contrappone a un piccolo gruppo di cellule germinali ( gonidia ), che però non è ancora sviluppato sotto forma di un organo delimitato . La presenza di testicoli (o, in linea di principio, di gonadi) non è una caratteristica fondamentale delle cellule multicellulari.
Nel caso dei bilateria compare per la prima volta un terzo cotiledone , il mesoderma , e quindi organi complessi. Tuttavia, in molti invertebrati la riproduzione sessuale è ancora combinata con la possibilità di riproduzione asessuata. C'è spesso un cambio generazionale qui , cioè quello sessuale segue un ciclo riproduttivo asessuato.
La differenziazione delle gonadi in testicoli e ovaie è un segno distintivo delle specie segregate. Finora non è stato chiarito se l'ermafroditismo o la segregazione di genere sia la caratteristica plesiomorfa della bilateria. Entrambe le gonadi emergono dallo stesso sistema nello sviluppo embrionale. In molti gruppi animali, nonostante questa separazione di genere, è possibile la riproduzione senza fecondazione ( partenogenesi ), che può essere vista come una forma ridotta di riproduzione sessuale. Gli animali maschi compaiono qui solo in casi eccezionali. La partenogenesi si trova in numerosi taxa , dai rotiferi ad alcune lucertole . Fino agli anfibi, durante l' ontogenesi si possono riscontrare anche forme ibride o un cambio di genere ( dicogamia ) . È possibile convertire le ovaie in testicoli ( proteroginia ) così come i testicoli in ovaie ( proterandria ).
Nella maggior parte degli animali, è geneticamente determinato se la disposizione inizialmente indifferente al sesso delle gonadi si traduce in un testicolo o in un'ovaia . Nei vermi e nelle mosche, il sesso è determinato dal rapporto tra cromosomi X e autosomi . Negli insetti che formano lo stato, i testicoli nascono nella prole da uova non fecondate e le ovaie negli animali con un insieme diploide di cromosomi, cioè da uova fecondate. Nei mammiferi, il sesso è determinato dal cromosoma Y. . Su questo cromosoma sessuale ( gonosoma ) si trova un gene ( regione determinante il sesso di Y , Sry ), che interagisce con i geni di altri cromosomi e (nell'uomo dalla 7a settimana dopo la fecondazione) porta alla formazione del fattore determinante il testicolo . Questo avvia lo sviluppo dei testicoli e quindi del sesso maschile in generale. Codifica una serie di fattori di trascrizione , le cosiddette proteine HMG ( proteine del gruppo ad alta mobilità ). Queste proteine hanno numerose altre funzioni in altri tessuti e gli esatti meccanismi coinvolti nella formazione dei testicoli sono attualmente oggetto di intense ricerche. Con l' espressione di Sry si differenziano le cellule del Sertoli, che tra l'altro producono l' ormone anti-Mülleriano e provocano quindi la regressione dei dotti Mülleriani . L'ulteriore sviluppo del testicolo e quello delle altre caratteristiche del sesso maschile è controllato dagli androgeni. Nel caso di alcuni gruppi animali, invece, il sesso è determinato da fattori ambientali. Nel caso di alcuni anfibi e di molti rettili ( tartarughe , alligatori ) , ad esempio, il sesso dipende dalla temperatura di incubazione. ( Vedi anche. Determinazione del sesso dipendente dalla temperatura )
In molti animali, le gonadi si sviluppano in stretta relazione con il sistema escretore ( rene primitivo , nefridi ), in particolare il tratto urinario è condiviso come sistema samenableitendes, motivo per cui entrambi i sistemi di organi nei vertebrati come apparato urinario e sessuale riassunti. Negli invertebrati, tuttavia, il luogo di origine, la posizione e i dotti sono molto diversi, per cui si presume che la riproduzione sessuale sia sorta più volte e indipendentemente l'una dall'altra nell'evoluzione . La complessità degli organi genitali non dipende dallo stadio di evoluzione, è molto alta nei platelminti, per esempio.
Embriologia dei vertebrati
Sviluppo testicolare nei vertebrati
|

Nell'embrione , i testicoli e le ovaie derivano dallo stesso sistema , la cosiddetta cresta genitale . Si forma nella zona del rene urnale e si estende inizialmente dal torace al lombo . Nella maggior parte dei vertebrati, solo la parte centrale di questo sistema allungato diventa la vera gonade, le sezioni rimanenti si sviluppano nei legamenti terminali . Sotto l'influenza di Sry, le cellule germinali primordiali del sacco vitellino migrate nella anlage gonadica (nell'uomo nella settimana embrionale 6a) e l' epitelio della cavità del corpo primitivo ( Celoma ) cresce simili a dita nel sistema come cosiddetti linee germinali primarie .
I cordoni germinali o testicolari penetrano nel sistema gonadico e crescono attorno alle cellule germinali primordiali. Le gonadi sono temporaneamente divise in corteccia e midollo, sebbene negli embrioni maschili solo il midollo si sviluppi nel testicolo, mentre la corteccia si ritira. Processi simili avvengono in individui geneticamente femminili, ma in seguito l' ovaio si forma dalla corteccia mentre il midollo degenera. Alla fine si perde la connessione tra i cordoni testicolari e la superficie. Dai cordoni testicolari, attraverso le cellule pre-Sertoli che esprimono Sry, si sviluppano le cellule Sertoli , che sono considerate gli organizzatori di un ulteriore sviluppo testicolare e quindi interagiscono con le cellule mioidi . Gli spermatogoni derivano dalle cellule sessuali primarie .
All'interno, i cordoni testicolari formano una rete di cordoni interconnessi, la successiva rete testicolare ( rete testis ). La rete testicolare si collega ad alcuni tubuli urnali che diventano i dotti efferenti della testa dell'epididimo . Il dotto urinario ( dotto di Wolff ) viene utilizzato anche come canale epididimale e dotto spermatico come percorso di drenaggio dello sperma. Il lume dei tubuli seminiferi non si sviluppa fino alla pubertà , negli anfibi dopo la metamorfosi , fino ad allora i cordoni testicolari sono solidi.
La tunica albuginea , la struttura del tessuto connettivo del testicolo e le cellule di Leydig derivano dalla parte mesodermica dell'ano testicolare . Le cellule di Leydig si trovano anche nel primo sviluppo testicolare; esprimono il fattore steroidogenico 1 (Sf1), e la loro produzione di testosterone determina significativamente lo sviluppo degli organi sessuali maschili.
Invecchiamento
Negli uomini - a differenza delle donne ( vedi menopausa ) - non c'è improvvisa cessazione della funzione delle gonadi ad un'età definita. Sia la produzione di ormoni che la maturazione delle cellule germinali sono potenzialmente preservate fino alla vecchiaia. La paternità , infatti, è stata documentata fino al decimo decennio di vita . In realtà, però, ci sono marcate differenze individuali e molti uomini prima o poi diventano sterili , cosa da non confondere con l' impotenza . Le cause esatte di queste differenze non sono note in dettaglio; si sospettano, tra gli altri, fattori vascolari . Dal punto di vista statistico, intorno alla quarta decade di vita in poi, l' involuzione dei testicoli , che progredisce molto lentamente, con diminuzione del peso, delle dimensioni e della produzione di spermatozoi. Per il singolo, invece, non ci sono quasi previsioni da fare.
Anche i cambiamenti strutturali sono quindi soggetti ad un ampio range, ma può essere considerato tipico un quadro misto di tubuli testicolari normali e chiaramente atrofici . Un marcato calo della produzione di testosterone osservato in alcuni uomini può portare a Climacterium virile con vampate di calore , mal di testa e altri sintomi .
Storia dell'esplorazione

Nell'antichità e nel Medioevo il testicolo era considerato solo una stazione di transito per il seme. Alkmaion von Kroton (inizi del V secolo a.C.) sospettava che il cervello fosse l'origine del seme, che raggiunge i testicoli attraverso i vasi sanguigni . Gli atomisti ( Anassagora , Democrito ) e Aristotele includevano il midollo spinale in questo percorso, Galeno (125-199) sospettava l'origine degli spermatozoi nei vasi sanguigni attraverso i quali raggiungono i testicoli. Queste idee persistettero nel Medioevo. I disegni anatomici di Leonardo da Vinci mostrano le connessioni tra il testicolo e i polmoni e il cervello, perché da Vinci sospettava l'origine del "potere spirituale" del seme nel cervello, mentre i testicoli forniscono solo la base materiale per gli "impulsi inferiori".
Esplorazione strutturale
Fu solo con l'inizio dell'Illuminismo nel XVII secolo che le idee sul seme maschile furono demistificate e fu riconosciuta la connessione diretta tra testicoli e riproduzione. La prima descrizione moderna della struttura del testicolo viene da Reinier de Graaf (1641–1673). Nathaniel Highmore descrisse nel 1651 il tessuto connettivo del testicolo ( Corpus Highmori ), che fu nominato nel 1830 da Astley Paston Cooper Mediastinum testis . Nel 1677, l' inventore del microscopio , Antoni van Leeuwenhoek , scoprì gli spermatozoi, che riteneva fossero organismi miniaturizzati e preformati ("animali-seme").

Con lo sviluppo delle tecniche istologiche , è stato possibile chiarire anche la struttura fine del testicolo. Nel 1841 l'anatomista svizzero Albert von Koelliker riconobbe per la prima volta la connessione diretta tra i tubuli testicolari e lo sperma e scoprì che gli spermatozoi in questi tubuli nascono come prodotti della differenziazione cellulare. Nel 1850, l'allievo di Kölliker Franz von Leydig descrisse per la prima volta le cellule intermedie ( cellule di Leydig ).
Nel 1865 Enrico Sertoli scoprì le cellule di supporto ( cellule di Sertoli ). Nel 1871 Victor Ebner riuscì a differenziare le cellule del Sertoli dagli spermatogoni e cinque anni dopo La Valette St. George coniò il termine "spermatogoni" e la classificazione dei singoli stadi di sviluppo degli spermatozoi, ancora oggi diffusa. Le cellule del Sertoli sono state a lungo considerate sincizie , non è stato fino al 1956 che Don W. Fawcett e Mario H. Burgos sono stati in grado di dimostrare che ogni cellula del Sertoli ha i propri confini cellulari.
Hugo Ribbert riconobbe già nel 1904 che il carminio somministrato nel sangue non entrava nel lume dei tubuli seminiferi e nella rete testicolare. Per molto tempo questa scoperta è stata trascurata, sebbene fosse la prima evidenza della barriera emato-testicolare. Questa conoscenza fu ripresa solo alla fine degli anni '50 e nel 1963 J. Brökelmann riuscì a dimostrare che le giunzioni strette delle cellule del Sertoli sono la base morfologica della barriera emato-testicolare. Paul J. Gardner e Edward A. Holyoke furono in grado di chiarire la struttura fine della barriera emato-testicolare un anno dopo.
Ormoni
Sebbene gli effetti della castrazione fossero noti da migliaia di anni, solo nel 1849 Arnold Adolph Berthold riuscì a dimostrare sperimentalmente la formazione di ormoni nei testicoli mediante trapianti di testicoli nei galli. In età molto avanzata, Charles-Édouard Brown-Séquard intraprese autoesperimenti alla fine del XIX secolo con il fluido dei testicoli di cani e porcellini d'India, a cui attribuì un potere ringiovanente e rinforzante, sebbene fosse più quantità di ormoni omeopatici che ha ottenuto in questo modo. All'inizio del XX secolo il trapianto di testicoli animali sotto la parete addominale era considerato un mezzo di ringiovanimento, in particolare i viennesi Robert Lichtenstern ed Eugen Steinach furono protagonisti di questo metodo. Steinach voleva ottenere questo processo di ringiovanimento legando i dotti deferenti (il suo paziente più famoso era Sigmund Freud ) e descriveva il trapianto di testicoli come una "terapia" per l' omosessualità . Dopo il 1945, questi controversi xenotrapianti passarono di moda.
Nel 1903, Pol Bouin e Paul Ancel pubblicarono per la prima volta la conoscenza che le cellule di Leydig sono il luogo in cui vengono prodotti gli ormoni sessuali maschili. Nel 1931 Adolf Butenandt e Kurt Tscherning isolarono l' androsterone (un metabolita del testosterone) dalle urine degli uomini, nel 1935 Ernst Laqueur riuscì ad isolare lui stesso il testosterone dai testicoli di toro e coniò anche il nome di questo ormone (da testis "testicoli" e " steroide ").
L'esistenza di ormoni non steroidei nei testicoli fu postulata già negli anni '20, ma fu solo nel 1932 che D. Roy McCullagh dimostrò la sua funzionalità nei ratti castrati e la chiamò inibina . Sebbene i test biologici per questo ormone siano stati sviluppati negli anni '60 , la sua esistenza è stata controversa per diversi decenni e non è stata ampiamente accettata fino al 1979. Nel 1984/85 furono rivelati la struttura e i sottotipi dell'ormone. Con la chiarificazione della sequenza del DNA che codifica per l'ormone , nel 1985 è stata riconosciuta l'affiliazione dell'inibina al gruppo dei fattori di crescita -trasformanti .
Mentre le relazioni tra LH e testosterone erano già note negli anni '60, la dipendenza dall'FSH delle cellule del Sertoli è stata dimostrata solo nel 1984 da Joanne M. Orth .
Differenziazione di genere
La base cromosomica della differenziazione dei sessi era già stata chiarita tra il 1910 e il 1916, principalmente attraverso il lavoro di Thomas Hunt Morgan sui moscerini della frutta , per i quali ricevette nel 1933 il Premio Nobel per la Fisiologia o la Medicina . Alfred Jost riconobbe nel 1947 che le gonadi sono determinate principalmente dal genere femminile e che il genere maschile dipende dal testosterone. Tuttavia, non è stato fino all'inizio degli anni '60 che il cromosoma Y è stato identificato come un fattore importante nei mammiferi. L'esatta posizione del gene per il fattore che determina i testicoli non è stata determinata fino al 1990; le sue diverse funzioni non sono ancora note in dettaglio e sono oggetto di ricerca attuale.
Disturbi e malattie dello sviluppo
Le lesioni al testicolo si verificano negli esseri umani come traumi contundenti , specialmente nelle arti marziali e nei combattimenti. Esiste il rischio di sanguinamento sotto la capsula testicolare ( ematocele ), che di solito richiede un trattamento chirurgico. Lesioni con apertura dello scroto (ferite da puntura e infilzamento, negli animali anche morsi, filo spinato, ecc.) possono causare infiammazioni testicolari (vedi sotto) o addirittura ascessi , nonché peritoniti dovute alla connessione aperta del processo vaginale al cavità addominale .
malformazioni
Un anorchy è l'assenza di entrambi i testicoli, se si forma solo testicolo, si parla di una monarchia . Circa il 5% dei bambini maschi operati per mancata discesa dei testicoli ha solo uno o nessun testicolo. Hanno spesso piccoli nodi di tessuto connettivo con cellule di Leydig sparse. Poiché un testicolo funzionalmente integro è assolutamente necessario per l'espressione del genere maschile, almeno un testicolo intatto deve essere presente nella fase embrionale, che può essere regredita in seguito.
In rari casi, a causa di disturbi dello sviluppo, le ovaie possono anche apparire oltre ai testicoli nell'uomo e in altri mammiferi ( ermafroditismo verus - ermafroditi "veri"; vedi anche intersessualità ). Con alcune malformazioni delle gonadi ( disgenesia gonadica ), i testicoli non si creano, rimangono sottosviluppati o contengono tessuto ovarico ( ovotestis ).
Uno sviluppo indesiderabile molto raro è la fusione splenogonadica con una connessione tra tessuto testicolare e milza .
Con disturbi della migrazione del testicolo ( Maldescensus testis ) possono verificarsi diverse anomalie di posizione . Il testicolo può rimanere nella cavità addominale ( criptorchidismo , " testicolo addominale"), rimanere incastrato nel canale inguinale ("testicolo inguinale", "scivolamento osseo") o prendere una strada sbagliata e, ad esempio, venire a giacere sotto la pelle dell'inguine o dell'interno coscia ( ectopia testicolare ). Il Maldescensus testis è una delle malformazioni più comuni nell'uomo e si manifesta nel 3-5% dei neonati e nel 33% dei prematuri; i criptorchidi si manifestano con analoga frequenza anche negli animali domestici e portano all'esclusione dalla riproduzione. Se il testicolo è posizionato in modo errato, lo sperma fertile non può formarsi a causa della sensibilità alla temperatura dell'epitelio germinale, ma viene mantenuta la produzione di androgeni. La mancata discesa per più di due anni può portare alla perdita di spermatogoni e quindi ad alterazioni irreversibili del testicolo.
La bassa statura ereditaria dei testicoli ( ipoplasia testicolare ) è relativamente comune negli animali domestici. L'ipoplasia testicolare può anche essere causata da disturbi cromosomici ( sindrome di Klinefelter ), infezioni o disturbi ormonali .
Un allargamento anormale dei testicoli è chiamato macroorchidia .
Infiammazione dei testicoli
L'infiammazione dei testicoli ( orchite ) può verificarsi con lesioni allo scroto con invasione di batteri o con alcune malattie infettive. L'orchite è una possibile complicanza della parotite , delle infezioni da virus Coxsackie e della varicella nell'uomo . Anche brucellosi e tubercolosi possono manifestarsi sui testicoli. Negli animali anche la tubercolosi e la brucellosi e la pseudotubercolosi ( pecora ), l' anemia infettiva equina e l' anemia infettiva felina ( gatto ) possono essere accompagnate da un'orchidite. L'infiammazione dei testicoli può portare al restringimento dei testicoli ( atrofia testicolare ) e alla sterilità , perché non si possono formare più spermatozoi ( aspermia ) o spermatozoi funzionanti.
Disturbi circolatori
Come un varicocele è chiamato vene varicose estensioni simili che in particolare le vene del lato sinistro del plesso pampiniforme riguardano nel funicolo spermatico. Un varicocele può portare a una produzione limitata di spermatozoi nel testicolo sullo stesso lato.
L' accumulo di liquido sieroso nelle guaine testicolari è chiamato idrocele o frattura dell'acqua. Inoltre, il fluido può anche raccogliersi nel cordone spermatico, che viene quindi indicato come idrocele funiculi spermatici .
Una torsione è una rotazione anormale del testicolo, in cui detto bloccaggio elicoidale del funicolo spermatico e delle vene drenanti può portare alla morte del testicolo. La torsione testicolare grave è un'emergenza molto dolorosa e ci si può aspettare un danno permanente al testicolo dopo appena due ore. L' appendice testicolare può anche sviluppare una cosiddetta torsione dell'idrato .
Disturbi circolatori con rischio di sviluppare necrosi si osservano anche in malattie dei vasi sanguigni come la porpora di Henoch-Schönlein , l' endangite obliterante e la panarterite nodosa nell'uomo, l' arterite nel cavallo e generalmente nella trombosi .
tumori
Un tumore testicolare è un allargamento patologico del testicolo. I tumori testicolari possono essere benigni o maligni .
Ingrandimenti testicolari per lo più innocui sono cisti . Sul testicolo possono svilupparsi due diversi tipi di cisti. Gli idroceli sono sporgenze della tunica vaginale testicolo che contengono un liquido limpido di colore ambrato. Sono causati da lesioni o infiammazioni. Gli spermatoceli originano dalla rete testis o dall'epididimo e contengono spermatozoi. I teratomi sono per lo più tumori benigni delle cellule germinali.
Una causa molto rara di una massa sul testicolo può essere la fusione splenogonadica .
I tumori testicolari maligni ( cancro testicolare ) si suddividono in degenerazioni delle cellule germinali ( tumori testicolari germinali : seminomi ) e non seminomi . La degenerazione delle cellule germinali è il cancro più comune negli uomini di età compresa tra 20 e 40 anni e rappresenta circa il 90% di tutti i tumori testicolari. Il principale fattore di rischio sono i testicoli che non sono migrati nello scroto.Il restante 10% è dovuto alla degenerazione tumorale di altre parti di tessuto ( tumore a cellule di Sertoli , tumore a cellule di Leydig , linfoma non-Hodgkin, ecc.).
Malfunzionamenti
Oltre alle malattie sopra citate , possono verificarsi anche sostanze chimiche che attraversano la barriera emato-testicolare come tossine ambientali (es. cadmio ), additivi ai materiali di confezionamento (es. ftalati , dietilesil adipato ), alcuni farmaci (es. furazolidone ) e ormoni ( vedi anche interferenti endocrini ) o radiazioni ionizzanti possono portare a una grave compromissione dell'epitelio dei tubuli seminiferi. Poiché la formazione degli spermatozoi è associata a tassi di divisione cellulare molto elevati ( mitosi , meiosi ), l'epitelio germinale è particolarmente sensibile alle tossine cellulari. Tale danno può portare a molteplici cambiamenti fino alla completa assenza di spermatozoi ( vedi anche analisi spermatica ).
La produzione inadeguata di androgeni è nota come ipogonadismo . Questo può essere congenito, secondario a malattie del testicolo o dovuto a un deficit di gonadotropine (es. ipofunzione dell'ipofisi , sindrome olfattogenitale ).
Criptorchidismo addominale in una pecora domestica ("cervo dei reni")

Ipoplasia testicolare in un gatto, a destra il testicolo normalmente sviluppato.

Varicocele distinto a sinistra.
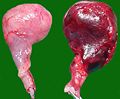
Testicoli del cane rimossi dopo la rotazione (a sinistra: organo sano)
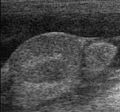
Ecografia : cane con criptorchidismo addominale e ascite (a sinistra: testicoli, a destra: epididimo)





visita medica
L' esame testicolare è un importante esame di base per l'uomo e gli animali con uno scroto. È qui che vengono verificate la presenza, le dimensioni, la posizione e la consistenza del testicolo. L' esame ecografico viene utilizzato principalmente come metodo di imaging . Il volume testicolare viene determinato mediante confronto con la cosiddetta catena di Prader o mediante misurazione ad ultrasuoni. La diafanoscopia ha ancora un certo valore nella diagnosi di idrocele . Una biopsia testicolare può essere eseguita per prelevare campioni di tessuto . Negli animali con testicoli situati nella cavità addominale, l' endoscopia viene utilizzata principalmente in aggiunta all'esame ecografico .
Un esame funzionale è la creazione di un'analisi del seme . Qui vengono valutati il numero, la forma e la mobilità dello sperma. La determinazione del livello di inibina B nel sangue viene utilizzata come marker per la funzione e la fertilità delle cellule di Sertoli, ma il suo valore informativo è controverso.
Il test di stimolazione cellulare di Leydig può essere utilizzato per rilevare il tessuto testicolare che non può essere rilevato utilizzando tecniche di imaging .
castrazione

La restrizione della funzione testicolare è chiamata castrazione. Può essere eseguita mediante rimozione chirurgica del testicolo ( orchiectomia ), legatura dei vasi testicolari ("castrazione senza sangue"), radiazioni o sostanze chimiche. Le castrazioni vengono eseguite principalmente nell'uomo per il cancro ai testicoli. I testicoli rimossi chirurgicamente sono solitamente sostituiti da una protesi testicolare per ragioni estetiche .
Come simbolo di impotenza, la castrazione svolge anche un ruolo nella mitologia di molte culture (vedi anche paura della castrazione ). Nella mitologia egizia, Horus strappa i testicoli al suo avversario Seth . Nella mitologia greca , Crono rimuove prima i testicoli di suo padre Urano e successivamente viene evirato dallo stesso figlio Zeus . Per rinunciare ai desideri mondani, era comune l'auto- evirazione dei Galloi (sacerdoti) nel culto di Cibele dei Frigi , che si diffuse anche nell'antica Grecia e a Roma, come nel caso degli Hijra in India . Nel Giudaismo, tuttavia , la castrazione, sia degli esseri umani e degli animali, è severamente vietato. Nel cristianesimo , anche la castrazione era disapprovata. Gli eunuchi non potevano essere ordinati sacerdoti, ma c'erano correnti in cui l'autocastrazione veniva eseguita come rituale (vedi Skopzen ).
Storicamente venivano castrati anche gli schiavi , i prigionieri di guerra, i cantori o le guardie degli harem (vedi Palazzo Eunuco ). La castrazione non giustificata dal punto di vista medico mirava in particolare alla prevenzione dei caratteri sessuali secondari (altezza della voce, comportamento sessuale) causati dal testosterone. I castrati erano popolari nella vita musicale europea nei secoli XVII e XVIII e spesso erano tenuti in grande considerazione. Senesino , Farinelli , Caffarelli e Antonio Bernacchi sono tra i più celebri castrati del Settecento .
Gli uomini castrati non possono riprodursi da soli. Simili ai chierici che si erano volontariamente astenuti , furono giudicati più affidabili e usati come funzionari e servitori in varie società. La castrazione volontaria dei reati sessuali è ancora un metodo terapeutico, anche se controverso, in Germania e in alcuni stati degli USA.
In medicina veterinaria la castrazione viene utilizzata, oltre alle indicazioni mediche (cancro ai testicoli, malattie della prostata e delle ghiandole anali ), soprattutto per evitare la prole, per migliorare la gestibilità degli animali da compagnia ( castrone , bue ), per aumentare le prestazioni di ingrasso e la qualità della carne, nei suini domestici anche per evitare l'"odore di cinghiale" delle carni effettuate. Le castrazioni sugli animali furono probabilmente effettuate già all'inizio del Neolitico . La castrazione è uno dei pochi prelievi di organi non indicati dal punto di vista medico ancora oggi consentiti in Germania ai sensi della legge sul benessere degli animali (§ 6), in animali molto giovani anche senza eliminare il dolore, il che, tuttavia, non è indiscusso.
Significato culturale e storico

Nella mitologia giapponese , i tanuki , demoni ( yōkai ) simili al cane procione , sono spesso rappresentati come simbolo di fortuna con i testicoli sovradimensionati. In Grecia , i genitali di animali, soprattutto i tori ( Taurobolium ), sono stati offerti come sacrificio. I testicoli erano considerati un simbolo di virilità e potenza creativa. Secondo Taylor, avevano un potere simbolico più forte del pene fino alla fine del XVI secolo .
botanica

Si pensava che le piante, che assomigliano ai genitali maschili nel loro aspetto, avessero un effetto afrodisiaco e di aumento della fertilità. Il filosofo greco Teofrasto von Eresos li chiamò Orchis , nome greco dei testicoli, per la somiglianza dei due bulbi dell'orchidea con i testicoli. Si dice che il loro consumo favorisca la nascita di un bambino ("orchidea"). Orchis in seguito diede il nome all'intera famiglia delle orchidee .
Il nome avocado deriva dalla parola indiana ahuacatl (testicolo), che si riferisce alla forma simile a un testicolo del frutto di questo albero.
nell'arte
In arte, a differenza del fallo , i testicoli non giocano un ruolo centrale al di fuori dell'erotismo e della pornografia . "Sangue e testicoli", un gioco di parole fonologico sull'ideologia del sangue e del suolo , è spesso usato in modo dispregiativo nella critica d'arte .
Uno dei personaggi del Tristan di Thomas Mann è Mr. Klöterjahn ("Klöten" è l' espressione basso-tedesca per i testicoli, simbolo di idoneità alla vita e vitalità). Nel romanzo del 2002 Sanningen om Sascha Knisch (Titolo inglese The Truth About Sascha Knisch. 2003) dell'autore svedese Aris Fioretos , i testicoli sono il leitmotiv . Il film tedesco Eierdiebe tratta il tema del cancro ai testicoli e della perdita di un testicolo.

Lo stemma della casata nobile italiana Colleoni di Bergamo mostra diverse coppie di testicoli ed è probabilmente un'allusione a coglione , un nome italiano per i testicoli.Una statua di Bartolomeo Colleoni con questo stemma sulla base prega il personaggio principale nella primo volume del romanzo in tre parti di Heinrich Mann Le dee o I tre romanzi della duchessa di Assy .
come cibo

I testicoli vengono trasformati come cibo in molte regioni. La Germania era l'unico paese dell'UE in cui i testicoli erano vietati come cibo. Secondo il regolamento UE n° 853/2004 del 29 aprile 2004, i testicoli sono gli unici organi genitali come cibo, tutti gli altri sono classificati come non idonei al consumo ( confische ).
varie
- L'americano Gregg Miller ha ricevuto il Premio Ig Nobel per la Medicina nel 2005 per lo sviluppo di protesi testicolari per cani castrati .
- Nel marzo 2006, la Volkswagen aveva pubblicizzato la Golf GTI su manifesti con "Turbo Cojones" negli Stati Uniti . In inglese, il termine cojones sta per coraggio e audacia, in spagnolo, dove cojones sta per "testicoli" e colloquialmente per "taglio" o "coraggio", la parola combinazione significa letteralmente "turbo testicoli". La promozione è stata ritirata dopo le proteste.
- Le ghiandole a forma di uovo di gallina (sacchi di ricino) sotto l'osso pubico del castoro erano anche conosciute come "testicoli" in passato.
letteratura
- AJP van den Brock: Gonadi e modalità di esecuzione. In: Bolk et al. (a cura di): Manuale di anatomia comparata dei vertebrati . Volume 6, Urban & Schwarzenberg, Berlino 1933, pp. 1-154.
- W. Busch, A. Holzmann (a cura di): Andrologia veterinaria. Schattauer, Stoccarda 2001, ISBN 3-7945-1955-8 .
- U. Gille: Organi riproduttivi maschili. In: F.-V. Salomone et al. (a cura di): Anatomia per la medicina veterinaria . Enke, Stoccarda 2004, ISBN 3-8304-1007-7 , pp. 389-403.
- R. Hautmann, H. Huland: Urologia . Springer, Berlino 2006, ISBN 3-540-29923-8 .
- H.-G. Liebich: Istologia funzionale dei mammiferi domestici. 4a edizione. Schattauer, Stoccarda 2003, ISBN 3-7945-2311-3 .
- JD Neill (a cura di): Knobil e la fisiologia della riproduzione di Neill. 3. Edizione. Academic Press, Amsterdam 2005, ISBN 0-12-515400-3 .
- PE Petrides: funzioni endocrine IV.Sistema ipotalamo-ipofisi e tessuti bersaglio. In: G. Löffler, PE Petrides (Ed.): Biochimica e patobiochimica. 7a edizione, Springer, Berlino 2003, ISBN 3-540-42295-1 , pp. 865-908.
- ONU Riede et al.: Sistema genitale maschile. In: ONU-N. Riede et al. (a cura di): Patologia generale e speciale . Thieme, Stoccarda 1989, ISBN 3-13-683302-3 , pp. 768-779.
- B. Vié: testicoli. Fête de paires, mitologia, lingerie, d'une curiosité culinaire, attributi du sujet, lexique. Edition de l'Epure, Paris 2005, ISBN 2-914480-58-X (numerose ricette, arricchite da informazioni culturali e storiche).
- R. Wehner, W. Gehring: Zoologia. 23a edizione, Thieme, Stoccarda 1995, ISBN 3-13-367423-4 .
- U. Welsch: Istologia da manuale di Sobotta. Urban & Fischer, Monaco di Baviera 2002, ISBN 3-437-42420-3 .
link internet
Evidenze individuali
- ↑ Walther Graumann: Anatomia compatta da manuale. Volume 3, Schattauer, Stoccarda 2004, ISBN 3-7945-2063-7 , pagina 265.
- ↑ H. Sosnik: Studi sulle dimensioni della gonade maschile umana nella biomorfosi, nell'intossicazione da alcol e nella cirrosi: una revisione e le proprie scoperte. In: Annuario morfologico di Gegenbaur. Vol. 134, No. 5, 1988, pp. 733-761, PMID 3224804 .
- ^ David AE Spalding: Balene della costa occidentale . Harbour Publishing, 1999, ISBN 1-55017-199-2 .
- ↑ I. Gerendai u a. .: innervazione e recettori serotoninergici del testicolo interagiscono con l'azione locale dell'interleuchina-1beta sulla steroidogenesi. In: Neuroscienze autonomiche. 7 luglio 2006, PMID 16829209 .
- ↑ I. Ducic, AL Dellon: Dolore testicolare dopo riparazione di ernia inguinale: un approccio alla resezione del ramo genitale del nervo genitofemorale. In: Journal of American College of Surgeons. Volume 198, n. 2, febbraio 2004, pp. 181-184, PMID 14759772 .
- ↑ FD Brown et al.: L'organo dell'offerente nel rospo Bufo marinus: Effetti dell'orchiectomia sulla morfologia e l'espressione del polipeptide associato alla lamina 2. In: Sviluppo Crescita e differenziazione. Volume 44, n. 6, dicembre 2002, pp. 527-535, PMID 12492511 .
- ↑ CF Farias: L'organo dell'offerente di Bufo ictericus: un'analisi di microscopia ottica ed elettronica. In: Micron. Volume 33, n. 7-8, 2002, pp. 673-679, PMID 12475564 .
- ↑ G. Michel: Sistema sessuale. In: F.-V. Salomon (a cura di): Manuale di anatomia del pollame . Fischer-Verlag, Stoccarda 1993, ISBN 3-334-60403-9 , pp. 197-226.
- ^ RP Amann, SS Howards: Produzione giornaliera di spermatozoi e riserve di spermatozoi nell'epididimo del maschio umano. In: Giornale di Urologia . Vol. 124, No. 2, agosto 1980, pp. 211-215, PMID 6772801 .
- ↑ C. Petersen, O. Soder: La cellula del sertoli - un bersaglio ormonale e un'infermiera "super" per le cellule germinali che determina le dimensioni dei testicoli. In: Hormone Research Volume 66, No. 4, 2006, pp. 153-161, PMID 16804315 (testo completo) .
- ↑ M. Fijak, A. Meinhardt: Il testicolo nel privilegio immunitario. In: Recensioni immunologiche . Volume 213, ottobre 2006, pp. 66-81, PMID 16972897 .
- ↑ RS Swerdloff et al.: La soppressione della spermatogenesi nell'uomo indotta dall'antagonista dell'ormone di rilascio della gonadotropina Nal-Glu e dal testosterone enantato (TE) è mantenuta dalla sola TE. In: Journal of Clinical Endocrinology and Metabolism . Volume 83, n. 10, ottobre 1998, pp. 3527-3533, PMID 9768659 .
- ↑ KL Matthiesson: effetti del testosterone e del levonorgestrel in combinazione con un inibitore della 5alfa-reduttasi o un antagonista dell'ormone di rilascio delle gonadotropine sulla spermatogenesi e sui livelli di steroidi intratesticolari negli uomini normali. In: The Journal of Clinical Endocrinology and Metabolism . Vol. 90, No. 10, ottobre 2005, pp. 5647-5655, PMID 16030154 .
- ↑ Peter Y. Liu et al.: Determinanti del tasso e dell'entità della soppressione spermatogena durante la contraccezione ormonale maschile: un'analisi integrata. In: The Journal of Clinical Endocrinology and Metabolism. Volume 93, n. 5, pp. 1774-1783, doi: 10.1210 / jc.2007-2768 .
- ↑ Improvac su vetpharm.uzh.ch
- ↑ a b c d C. Tilmann, B. Capel: vie cellulari e molecolari che regolano la determinazione del sesso dei mammiferi. In: Recenti progressi nella ricerca sugli ormoni. Volume 57, 2002, pp. 1-18.
- ↑ C. Dournon et al.: Inversione del sesso della temperatura negli anfibi e nei rettili. In: International Journal of Dev Biology. Volume 34, numero 1 marzo 1990, pp. 81-92. PMID 2393628
- ^ E. Nieschlag et al.: Funzioni riproduttive nei giovani padri e nonni. In: Journal of Clinical Endocrinology and Metabolism . Vol. 55, 1982, pp. 676-681.
- ↑ H. Bürgi, C. Hedinger: Cambiamenti istologici testicolari nella vecchiaia. In: settimanale medico svizzero. Volume 47, 1959, pp. 1236-1239.
- ↑ A. von Kolliker: Contributi alla conoscenza dei rapporti sessuali e del liquido seminale degli invertebrati, insieme ad un esperimento sulla natura e sul significato dei cosiddetti animali seme . Berlino 1841.
- ↑ AHJ La Valette St. George: Sulla genesi dei corpi seme. In: Archivi di anatomia microscopica. Volume 12, 1876, pp. 797-825.
- ^ J. Brökelmann: Struttura fine delle cellule germinali e delle cellule di Sertoli durante il ciclo dell'epitelio seminifero nel ratto. In: Ricerca cellulare e anatomia microscopica. Vol. 59, 1963, pp. 820-850, PMID 14015736 .
- ^ PJ Gardner, EA Holyoke: struttura fine del tubulo seminifero del topo svizzero. I. La membrana limitante, la cellula di Sertoli, gli spermatogoni e gli spermatociti. In: Scheda Anatomica . Volume 150, dicembre 1964, pp. 391-404, PMID 14248309 .
- ↑ AA Berthold: Trapianto di testicoli. In: Archivi di anatomia e fisiologia. Reparto Fisiologico . Volume 16, 1849, pp. 42-46.
- ↑ F. Mildenberger: Ringiovanimento e "guarigione" dell'omosessualità. Eugen Steinach ai suoi tempi. In: Ricerca sul sesso. Volume 15, 2002, pp. 302-322.
- ^ P. Bouin, P. Ancel: Recherches sur les Cellules interstitielles du testicule chez les mammifères. In: Archives de zoologie expérimentale et générale. Serie 4, Volume 1, 1903, pp. 437-523.
- ↑ K. David et al.: Informazioni sull'ormone cristallino dei testicoli (testosterone). In: Giornale di chimica fisiologica . Volume 233, 1935, pp. 281-282.
- ↑ A. Lampel: Commento. In: Urologia attuale. Volume 35, 2004, pp. 6-8. (testo completo) ( Memento del 14 novembre 2007 in Internet Archive )
- ↑ Axel Wehrend: sintomi chiave di ginecologia e ostetricia nei cani. Enke, Stoccarda 2010, ISBN 978-3-8304-1076-8 , pagina 57.
- ^ A b c G. Taylor: Castrazione: una storia abbreviata della virilità occidentale. Routledge, 2002, ISBN 0-415-93881-3 .
- ↑ W. Bittorf: sangue e testicoli . In: Der Spiegel . No. 44 , 1976, pp. 228-232 (in linea - 25 ottobre 1976 ).
- ^ Peter Philipp Riedl: Quadri d'epoca - tipologie di artisti. Contributi a bozze tradizionali in letteratura e scienza dal 1860 al 1930. Vittorio Klostermann, 2005, ISBN 3-465-03410-4 , ( p. 570 ).
- ↑ Il regolamento (CE) n 853/2004 del Parlamento Europeo e del Consiglio. ( europa.eu )
- ↑ Miriam Jordan: Good Taste Lost In Ad Translation . Su: items.sun-sentinel.com dal 19 marzo 2006 (inglese).
- ↑ 'Turbo-Cojones', il eslogan de Volkswagen que en molesta EEUU . Su: elmundo.es del 21 marzo 2006 (spagnolo).
- ↑ Foto del manifesto. Su: secure.flickr.com dal 20 marzo 2006.
- ↑ Dieter Lehmann: Due ricettari medici del XV secolo dall'Alto Reno . Parte I: Testo e Glossario . Horst Wellm, Pattensen/Han. 1985; ora (= ricerca storico-medica di Würzburg. Volume 34). Königshausen & Neumann, Würzburg, ISBN 3-921456-63-0 , pagina 194.